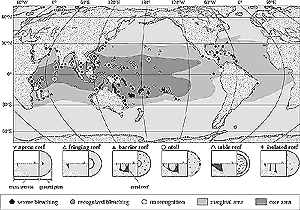
Figure 19-3: Bleaching of coral reefs reported from 1997 to 1998 (Mimura and Harasawa, 2000).
Coral reefs are restricted to narrow latitudinal, horizontal, and vertical ranges along the tropical continental shelves. Their contribution to global coastal biodiversity is disproportionate to their spatial extent: Although they cover less than 1% of the world's oceans, they are inhabited by one-third of globally known marine species (Reaka-Kudla, 1996). Coral reefs have far-reaching effects; they are nurseries for many ocean fish species, and they protect coastlines from wave impact and erosion (see Section 11.2.4.3). Thus, fisheries, tourism, infrastructures, societies, and cultures depend on the well-being of this unique entity that is impacted by increased temperature, atmospheric CO2, and sea level, synergistically combined with anthropogenic stresses that are independent of climate change.
Many reef-building coral species already live close to their upper thermal limit (see Section 6.4.5). If they are exposed to moderate increases (1-2°C) in water temperature, they become stressed and experience bleaching (Goreau et al., 1998; Hoegh-Guldberg, 1999). The increasing frequency of coral bleaching events during the past decade provides a reason for concern for this warming-induced impact. For example, 50-90% bleaching-induced mortality in the Indian Ocean reefs was associated with a 2-6°C above-normal sea-surface maximum triggered by El Niño during 1997-1998; several other severe bleaching events occurred in the 1982-1983 and 1987 El Niño years (Glynn, 1991; Wilkinson et al., 1999; see Figure 19-3).
Figure 19-3: Bleaching of coral reefs reported from 1997 to 1998 (Mimura and Harasawa, 2000). |
Defining the upper thermal thresholds of corals and using global warming scenarios, Hoegh-Guldberg (1999) found that the frequency of bleaching is expected to rise until they become annual events in most oceans (at about a 1°C warming). In some areas, bleaching events would happen more frequently as early as 2020 (with less than 0.5°C warming); within the next 30-50 years, bleaching could be triggered by normal seasonal changes in seawater temperature, and most regions may experience severe bleaching conditions every year. This trend exceeds the frequency at which corals can effectively recover from bleaching-related mortality (Hoegh-Guldberg, 1999).
Besides the detrimental effect of temperature rise, increased atmospheric CO2 concentrations reduce coral calcification rates (Gattuso et al., 1999; Kleypas et al., 1999; see Section 6.4.5), which already might have decreased and could decrease an additional 10-30% by 2100 (see Chapter 6). A 10-20% decrease in calcium carbonate production may impair expansion of coral reefs into higher latitudes as a response to predicted increasing SST (Kleypas et al., 1999). Healthy reef flats may benefit from projected increased sea level because they would be able to keep up with the projected rise in sea level. However, any increase in the frequency of El Niño and other ocean-atmosphere interactions, such as Indian Ocean dipole events, will lead to regular and prolonged sea-level depressions (10-40 cm) in the western Pacific and eastern Indian Ocean, with adverse effects on shallow reefs in their regions (see Chapter 6).
Mangrove ecosystems resemble coral reefs in that they have a narrow global distribution: They cover 11,500 km2 in all of Australia, and the largest mangrove forest in the world—the Sundarbans of Bangladesh and India—covers 6,000 km2 (see Section 11.2.4.1). Yet they have a rich biodiversity and a significant effect on adjacent and distant systems. Mangroves are made up of salt-adapted, intertidal evergreen trees on low-energy, sedimentary tropical shorelines, extending landward in lagoons, estuary margins, and tidal rivers (see Sections 6.4.4 and 11.2.4.1).
Mangrove ecosystems are highly vulnerable to sea-level rise induced by climate change, which will change the salinity distribution and inundate mangroves. For example, a 45-cm rise could inundate 75% of the Sundarbans (see Section 11.4.1); a 1-m rise will completely inundate the Sundarbans (see Section 11.2.1.6). In addition, redistribution of species whose habitats will be affected by inundation may be impaired because migration, especially to the north, will be blocked by human settlements. Loss of productivity, species, and ecosystem goods and services therefore is expected. Climate change effects will be further exacerbated, and vulnerability to climate change will increase human-induced damage. For example, between 56 and 75% of different Asian mangrove forests have been lost during the 20th century because of overexploitation and replacement by aquaculture installations (see Section 11.1.3.1). Like the Sundarbans, the Port Royal mangrove wetland in Jamaica may completely collapse with a 1-m sea-level rise (see Section 17.2.4.2).
On the other hand, although mangroves are vulnerable, some may be adaptable to climate change because they could survive in areas where vertical accretion equals sea-level rise. Because sediment flux determines mangrove response to sea-level rise and fluxes vary between regions and locations, the fate of the world's mangrove ecosystems will not be uniform (see Section 6.4.4). Yet even where accretion will offset sea-level rise, any infrastructure will limit the potential for landward migration of coastal mangrove species and habitats (see Section 6.2). Thus, at least some if not many of the world's mangrove ecosystems are unique and threatened entities.
|
Other reports in this collection |
|